In this brief note we add other elements, namely the
probability that a cell can mutate and that as it mutates the factors related
to the propagation model may also change. We calculate a similar diffusion
equation now for the average number of malignant cells by region and by type.
That is we demonstrate in the following graphic summary:

1. The standard diffusion-flow-proliferation model applies
on a per-region and per cell type basis. This means that the constants we have
developed previously will depend on the specific cell type as well, namely how
many mutations have occurred.
2. That we know there are multiple mutations in cancer
cells. Some may have a few and are indolent and others may have many and be
aggressive. We develop a Markov model for such cell progression.
3. We combine the three element spatio-temporal model with
the Markov cell mutation model and this allows us to determine the average
number of cells of a specific type in any part of the body at any point in
time.
4. We then discuss how one may use this model for prognostic
and therapeutic purposes.
The main observation in this brief section is that the
average number of malignant cells of a specific mutation state can be
determined by the following:

In this equation the n is an NX1 vector of average numbers
in spatio-temporal dependent values of each of N possible mutations and the L
value is the spatio-temporal dependent operator matrix and Λ is a matrix
describing the Markov transition probabilities between mutations.
It should be clear that we can measure all of the constants
involved and thus determine the result. As a counter-distinction we can measure
the n values and mutation states and determine the constants.
The expanded model considers the issue diagrammed below:
The next issue is the ability to determine what the factors
are in the specific model, namely the values of the constants, and secondly the
validation of the model itself.
Thus there are two dimensions of issues here:
1. Model Identification and Validation: In previous work we
referred to this as the Observability problem. Namely if we have a model and we
can identify the required parameters, then can this model be used to determine
the end state which will be attained. This is the prognostic problem.
2. Model Utilization: As with the previous cases, if we have
this model, and we have identified the constants, can we determine actions which
may be taken to control the end state of the system? This is the
Controllability problem. It states that perhaps having such a model we can
determine methods and means to drive the system, in this case the average
number of malignant cells of genotype say G, to a new end state, one where we
have reduced the number of bad cells to a de minimis level. This is the
therapeutic problem.
There also is a third element:
3. Identification: In both of the two previous issues we
assumed that there existed a method by which we could determine the constants
of diffusion et al and furthermore that we could ascertain the list of possible
mutations, and also their Markov transition probabilities. This may be
accomplished in two ways. First, we can accomplish this by in vitro studies.
Second, we can achieve this by using the model itself in a classic system
identification model with in vivo analyses.
Thus the analysis contained herein is an initiation of what
appears to be an innovative way to look at cancer. There have been many studies
in more specific and segmented areas but there has not to my knowledge been a
study that has examined cancer in such a broad and overarching manner. In
essence we have included all of the variables that one may ask for.
To better understand we depict the progression of prostate
cancer below.
Step 1: Benign State, here we have five segments; prostate,
two tissue-blood barriers, blood, and bone.
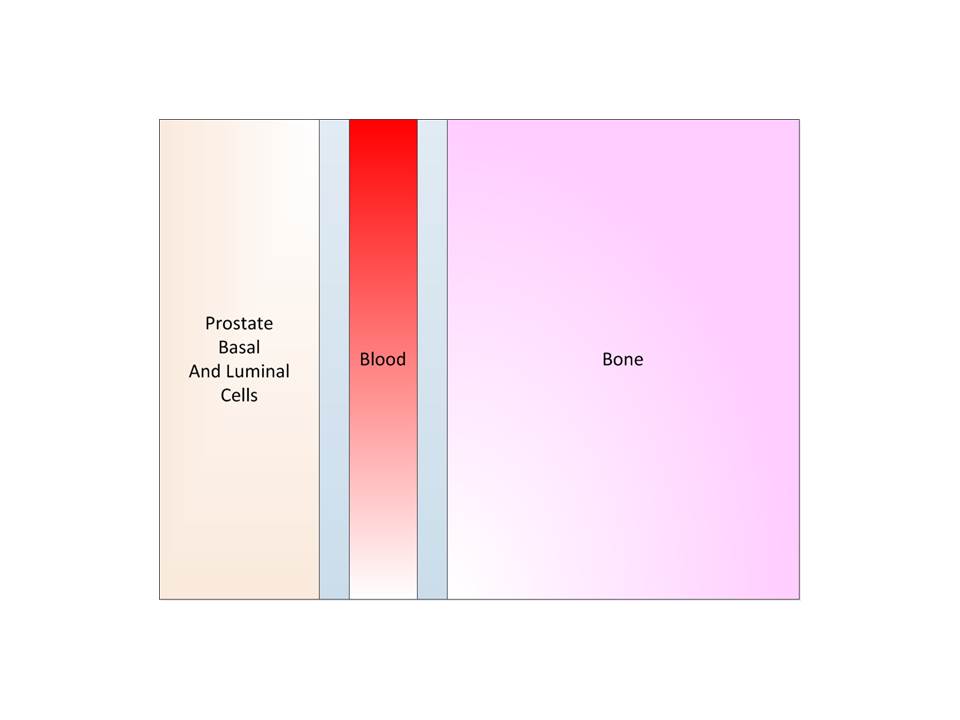
Step 2: We have the beginning of a cancer due to some
mutation of the basal or luminal cells. The cancer proliferates and diffuses.
It is still localized here.


Step 4: The blood barrier is crossed, and we assume by
diffusion. Across this barrier there is no proliferation or flow, just
diffusion.

Step 5: The blood barrier is crossed and the cell is now in
the blood stream. Here we have flow but no diffusion and no proliferation.
Step 6: The blood barrier is crossed again as discussed
above.

Step 7: Metastasis is complete by having the new malignant
cells in the bone and proliferation and diffusion predominate.

The above steps are common is almost all cancers. The
assumptions here are:
1. The same malignant cell moves across the body.
2. Each separate area, in this case five, has constant
diffusion, flow and proliferation constants.
3. That we can then measure the number of cells from this
deterministic model.
In the case where they are uniform constants we can solve
the equation. In the case where they are uniform constants across uniform
spatial domains then we can also solve the equations evoking boundary
conditions.
We now want to expand this model to include multiple
malignant cell types. Also we want to include their stochastic dynamics as
well.
Consider a cell with five possible mutations. We show the
genes below. The call may begin with one mutation and then move to a second and
so forth. Each path is assumed to be possible and the results of each path are
different.

Now we can consider a model for the above simple example. We
have 5 possible mutations and they may occur in any order. We assume they occur
one at a time. We can identify any number of cells as n(x,t), as the number of
cells after one mutation at location x and at time t, of mutation k.
Now we have the following observations:
1. At mutation 1 we have 5 possible cell mutants.
Furthermore each may be considered a cancer cell and the growth, diffusion and
flow are as described above. Some of the mutations may be indolent and some
aggressive.
2. At mutation 2 we have 5*4 possible cells. The question is
that some are say PTEN then cMyc or cMyc then PTEN. Are they the same, and this
means the difference between perturbation and combination? Are they distinct by
have been ordered differently or are they the same? If it is a combination we
have 10 instead of 20 different mutations.
3. At mutation 3 we have 5*4*3 and at 4 we have 5*4*3*2 or
120 permutations.
4. At any location we may have any one or a combination of
these mutation types. There are two factors driving their number:
a. A single type will have growth, dispersion and movement
dynamics with the above mentioned model but each mutation will respond
differently since their coefficients will be different. Some may grow faster
and some may diffuse faster. There is no a priori ranking of the coefficients.
b. The surrounding mutant types will also tend to mitigate
growth.
For example consider the following three gene mutation case:

We can thus make several important observations regarding
this model.
1. Prognostic and Therapeutic: We can determine the
transitions and the factors related to diffusion, flow and growth. Thus we can
use the result as a powerful one for prognostic and therapeutic results. As we
had indicated earlier, the Observability and Controllability issues are
essentially Prognostic and Therapeutic respectively.
2. Variances: The results are for the average. We can
determine the results for the variances as well. We have examined the variances
on the averages and they are somewhat complex and we do not believe that they
lend significant additional information at this time.
3. Solutions: The solutions to these equations are readily
obtained using standard techniques. They can, in addition, be determined in
closed form results.


