In a recent piece in Eureka the article refers to studies which present concerns regarding the migration of species across an open seas in the Arctic.
They state:
For the first time in roughly 2 million years, melting Arctic sea ice
is connecting the north Pacific and north Atlantic oceans. The newly
opened passages leave both coasts and Arctic waters vulnerable to a
large wave of invasive species, biologists from the Smithsonian
Environmental Research Center assert in a commentary published May 28 in
Nature Climate Change. Two new shipping routes have opened in the Arctic: the Northwest
Passage through Canada, and the Northern Sea Route, a 3000-mile stretch
along the coasts of Russia and Norway connecting the Barents and Bering
seas. While new opportunities for tapping Arctic natural resources and
interoceanic trade are high, commercial ships often inadvertently carry
invasive species. Organisms from previous ports can cling to the
undersides of their hulls or be pumped in the enormous tanks of ballast
water inside their hulls. Now that climate change has given ships a new,
shorter way to cross between oceans, the risks of new invasions are
escalating.
Now some brief thoughts. First if some higher species decided Homo sapiens should not cross from Asia to the Americas I wonder what would have happened here? Second, an open Arctic passage has been sought for centuries and perhaps this may lead to substantial changes in trade; for better or worse. Third, we have argued that putting fiber across the Arctic, North America and Russia, would be highly productive. This may open up that opportunity.
Wednesday, May 28, 2014
Monday, May 26, 2014
Memorial Day: Remembering

Tuesday, May 20, 2014
Re-admissions: An Interesting Difference
The issue of re-admissions has been a major concern for increased Health Care costs in hospitals. In a recent study by H-CUP they reported on a cross section.

The above is the summary of Re-admissions for Medicare. No real surprises here. In contrast look at Medicaid below:
 They are almost dominated by psychiatric causes. Also for Private Insurance we have:
They are almost dominated by psychiatric causes. Also for Private Insurance we have:
 Again a somewhat similar profile.
Again a somewhat similar profile.
This says a great deal about what we expect in the ACA. Two separate segments.
This says a great deal about what we expect in the ACA. Two separate segments.
FEDs Balance Sheet May 2014

The following chart depicts the key drivers.

It will ne necessary to watch this continued explosion. It is well over $5 trillion when before the Crash it was only $800 billion. We will see an ten fold increase by 2016.
Budget and Actuals on Discretionary Spending


Now the following is the kicker:

Earth Invades Mars!

As Nature states:
Dozens of microbial species may have accompanied the Curiosity rover to Mars, where it landed in August 2012. The stowaways withstood spacecraft cleaning methods before the rover's launch, although no one knows for sure whether the bacteria survived the inter-planetary ride. A study that identified 377 strains found that a surprising number resist extreme temperatures and damage caused by ultraviolet-C radiation, the most potentially harmful type. The results, presented today at the annual meeting of the American Society for Microbiology, are a first step towards elucidating how certain bacteria might survive decontamination and space flight.
After all the care we take in not contaminating our water etc we allow NASA to act with abandon in sending some of the most virulent of Earth's species to our nearest neighbor. Where is the EPA on this one?
The article continues:
Although spacecraft go through multiple cleaning steps to ensure that they bear no biological contaminants, previous reports suggest that Curiosity project developers did not follow these planetary protection protocols to the letter. The regulations are a safeguard; whether microbes can tolerate conditions on the surface of Mars is still unknown. “We don’t know yet if there’s really a threat,” says Smith. “Until we know, it’s important to take a precautionary approach.”
That is rather neglectful one could say. Next we will be sending our TV and movie products to totally contaminate them!
Monday, May 19, 2014
Obesity: A Scale versus the Genes
There is a continual debate over the idea that:
Input-Output=Net Accumulation
This is always true, by definition.
However we should always examine what we mean by each term.
Let us first examine Input.
Input= the net amount of kcal an individual obtains by the
consumption of a certain type of food
Now that will depend on the food and the individual. Certain
people have enzymes, products of genes, that convert the consumed more efficiently,
and they also have epigenetic factors that enhance conversion of the consumed
food into products that can be used or stored. Thus Input for one person
consuming the same food as another is NOT the same. Genes play a part in how
the physical raw input is processed in the individual to stored and used Input
in the body. Thus the net Input, that which is used and/or stored, may vary in
some statistical manner between humans.
For example certain southwest Indian tribes have genes that
allow highly efficient absorption meaning that they can live on poor quality
food but give rich foods they explode in weight and Type 2 Diabetes. Another
example, epigenetic, is from Netherlands in WW II where in 1944 mothers were
starved by Nazis and children born had epigenetic marks that allowed them to
survive on low caloric intake by up-regulating certain genes. Thus genetic and
epigenetic factors affect the conversion of Input and the net result can vary.
The epigenetic factors can be passed down to children and in some cases to
grandchildren. Thus environment via epigenetic changes can affect the genetic
makeup.
Having said this, one can develop a distribution of net
Input as a function of Gross Input, namely consumed food, and see that across a
large population there is, one can assume, a Gaussian like distribution. Some
people, most if you will, have a conversion rate of say 1, others may have a larger
or smaller conversion rate. It does not appear that a great deal has been done
examining this factor.
Output is basal metabolism rate and other factors whereby we
burn calories. That also varies dramatically. You walk 5 miles and I walk 5
miles and we each urn a different amount. Again Output may vary from individual
to individual. I may burn 100 kcal per mile walked and another may bur 75 or
125 kcal. Why the difference? Again it may be an amalgam of genetic or
epigenetic. It may also be the way one walks. Therefore Output like Input has
some form of distribution across large populations.
The problem thus is neither Input nor Output, it is Net
Accumulation. Thus for zero Net Accumulation we each must understand our
balance. Now that also may change as we may face different challenges.
Humans have the ability to use their intellect to measure a
set point, namely a scale and to measure their weight. Secondly humans have the
will power and intellectual capability to retain that set point.
Other animals spend all day hunting for feed and consuming
generally low calorie food, and somehow maintain a balance. Rarely do we see a
fat squirrel
The conclusions are:
1. Environment does affect what we see as net Input and net
Output and thus net Accumulation
2. Genetics and Epigenetics affect what each individual does
in terms of conversions
3. However as humans we have the ability/intelligence to
measure the set point and the will to maintain it.
4. One size fits all does not work. Each human is different
in how they convert Gross Input to Net Input and Gross Output to Net Output.
Thus the only true measure is the scale, namely weighing oneself and balancing
Input and Output to keep the scale at the required set point.
Thus when writes in the NY
Times and other journals of note speak of diet and weight control we hear
them state[1]:
FOR most of the last century, our
understanding of the cause of obesity has been based on immutable physical law.
Specifically, it’s the first law of thermodynamics, which dictates that energy
can neither be created nor destroyed. When it comes to body weight, this means
that calorie intake minus calorie expenditure equals calories stored.
Surrounded by tempting foods, we overeat, consuming more calories than we can
burn off, and the excess is deposited as fat. The simple solution is to exert
willpower and eat less. The problem is that this advice doesn’t work, at least
not for most people over the long term. In other words, your New Year’s
resolution to lose weight probably won’t last through the spring, let alone
affect how you look in a swimsuit in July.
All too often they oversimplify all factors. The basic law
holds, it is a definition, a tautology. The problem is the conversion rates
from Gross to Net, a factor that the authors above seem to struggle with but in
apparent ignorance. Basically they argue about some set point theory. Namely
humans eat until the set point and then stop. Eat too much, get fat, the set
point increases and the system is unstable. Fat Squirrels! However this totally
ignores the scale. If the goal is a BMI of 24.0-25.0 worst case, then just use
the scale, reduce food intake by the use of the will until the scale falls back
to the right range, and forget these putative set points, namely blaming some
other factor. Take responsibility.
The authors continue:
If this hypothesis turns out to
be correct, it will have immediate implications for public health. It would
mean that the decades-long focus on calorie restriction was destined to fail
for most people. Information about calorie content would remain relevant, not
as a strategy for weight loss, but rather to help people avoid eating too much
highly processed food loaded with rapidly digesting carbohydrates. But obesity
treatment would more appropriately focus on diet quality rather than calorie
quantity.
Obesity treatment is a non-issue by using a scale. Yes
indeed, forget the calories, measure the pounds. Scales are cheap!
Alan Turing: Calico Cats, Zebras, and Daylilies
Turing in 1954 wrote his paper on Tessellation, the patterns we often see on animals and plants. The question is; is Turing’s approach the right way to understand these patterns? Namely Turing proposed a hypothetical method whereby cells communicate with one another and that this communications is akin to flows of some, as of 1954, yet to be defined chemical substance or substances. Depending on the concentrations of these substances the cells then turned, for example, black or white, as in a zebra, and that this flow being coordinated in some manner yielded a pattern, not just a mass of black and white hairs.
Turing hypothesized, for example, that there may be two
controlling molecules in varying densities and if one molecule was denser than
the other it would turn on white and otherwise it would turn on black. But
Turing said more, that the flow of the molecular density was not just random
but that cells somehow participated in a distributed manner so that the
densities flowed as waves, with peaks and valleys. Thus the Zebra stripes were
a reflection of this flow. When the black molecule was at a high the hairs were
black and when the white ones were high the hairs were white. The net result
looked like waves of white and black.

Now a second model that has become of recent popularity is
the explanation for the Calico Cat.

The explanation for this is dramatically different. Here
Calico Cats are all female. The way it works is the epigenetic silencing of one
of the X chromosomes. This allegedly is totally done at random. As seen above
this of course is hardly the case. If every hair cell were random then we would
expect to have a blend of two or more colors and not the patterns that we have
above. This means that if this is epigenetic that spatially there is some
mechanism that is not totally random. There is some form of cell to cell memory
and cell to cell thresholding. Namely the hair stays black, brown or white for
some spatial period and then switches to the other state. That is the Turing Tessellation
effect. What then does that?
Now consider a third example; the daylily. We show a typical
example below. Here we have an eyezone, the dark red around the milled and we
even have red on the edges. This is again a tessellation type as described by
Turing. There are areas where there is dark red and areas where there is light
red or pink. Is this epigenetic, genetic, or a Turing tessellation.

In fact daylilies can show dramatic patterning as they get
more sophisticated. The above is a simple form of patterning, a wave of dark
and light red.
Thus what enables these patterns? In epigenetics of cats, it
is the turning on and off of one X chromosome, but not really randomly, so
there must be some mechanism that selects which one gets wrapped in an lncRNA
and which does not. What is that activator? Not yet known.
In daylilies we know that color is driven by anthocyanin
production. More of one and we get one color and more of the other another
color. We also know that certain proteins, gene products, act as catalysts
facilitating one anthocyanin path or another. Thus if one gene is producing
then it may drive up pone anthocyanin or another. What turns these genes on and
off? Perhaps epigenetic methylation as we see in many other examples. But that
begs the question of what causes the methylation? It appears to be a pattern
like that of the Calico Cat.
Thus the Turing Tessellation is a process that explains
these patterns but the facilitator of that process, the extracellular or
intercellular molecule is not known. Thus we have an interesting area of
exploration. In addition we see similar effects in the field of metastatic
cancers, where we get clusters of metastatic cells, and not just random
aberrant one.
Friday, May 16, 2014
Obesity and Willpower
Does the law of mass balance really apply to human obesity? Namely does Input Less Output Equal Net Accumulation really work?
In a recent NEJM article one is asked to wonder. Everywhere else in the universe it applies but not for some strange reason in human obesity. The author states:
These studies, of course, reinforce what some physician–researchers have been insisting for more than a century: that obesity is innate, that weight regulation is not governed by a uniform tally of “calories in–calories out,” and to quote ..., that “there is a biochemical or basic biological element in what it is that we call `willpower.'” The views of many Americans notwithstanding, weight is clearly far from being entirely within an individual's control. Genetic predispositions, in tandem with the development of food environments that facilitate overeating and built environments requiring minimal energy expenditure, may help explain why so many Americans are obese today.
Unfortunately the statement is false, almost in toto. Yes, each person has a different set point, namely the Output level. Some people burn 1500 kcal per day and others 2200 kcal per day. There is no meter telling us that. In addition that Output number does depend on a lot of factors, stress, ages, health. But if Input exceeds Output then Net Accumulation occurs. The opposite is also true. Weight management is often a sophisticated balance, but the true meter is a scale. If one targets say a BMI of 24.0 then one just manages Input to ensure the 24.0 number is reached and maintained.
It is always nice to have that extra piece of pie, those cookies, that extra beer, I guess, but that is input. Yes there are people who eat anything and never gain a pound and those of us who measure calories each time we come near food.
As the author starts out:
The obese lack willpower; they overeat and underexercise — or so believe a majority of Americans. A 2012 online poll of 1143 adults conducted by Reuters and the market research firm Ipsos found that 61% of U.S. adults believed that “personal choices about eating and exercise” were responsible for the obesity epidemic. A majority of Americans, it seems, remain unaware of or unconvinced by scientific research suggesting that “personal choices” may not account for all cases of obesity.
No one doubts that there are a small few for whom weight control is highly problematic. No matter how low they go their Output is set well below that point. The few calories expended in exercise will never increase the output above the minimal input. But that does not account for the near global explosion of obesity. There was not a genetic change in humanity over the past thirty years to account for this. There is no epigenetic hyper/hypo methylation setting and resetting genes, causing massive reductions in basal Output. Input has increased by "choice" and all attempts to rationalize it as some effect out beyond human control is both wrong and harmful.
The law of mass balance applies. No one has ever rejected this. If it works in any chemical plant, and the human body is one, then it works in us. Remember the best meter is that scale and the best control mechanism is to not eat that extra candy bar or drink that extra soft drink. It is truly a shame that some physicians cannot call it for what it is. Oh, it appears as if the author is from the History Department at Harvard...
In a recent NEJM article one is asked to wonder. Everywhere else in the universe it applies but not for some strange reason in human obesity. The author states:
These studies, of course, reinforce what some physician–researchers have been insisting for more than a century: that obesity is innate, that weight regulation is not governed by a uniform tally of “calories in–calories out,” and to quote ..., that “there is a biochemical or basic biological element in what it is that we call `willpower.'” The views of many Americans notwithstanding, weight is clearly far from being entirely within an individual's control. Genetic predispositions, in tandem with the development of food environments that facilitate overeating and built environments requiring minimal energy expenditure, may help explain why so many Americans are obese today.
Unfortunately the statement is false, almost in toto. Yes, each person has a different set point, namely the Output level. Some people burn 1500 kcal per day and others 2200 kcal per day. There is no meter telling us that. In addition that Output number does depend on a lot of factors, stress, ages, health. But if Input exceeds Output then Net Accumulation occurs. The opposite is also true. Weight management is often a sophisticated balance, but the true meter is a scale. If one targets say a BMI of 24.0 then one just manages Input to ensure the 24.0 number is reached and maintained.
It is always nice to have that extra piece of pie, those cookies, that extra beer, I guess, but that is input. Yes there are people who eat anything and never gain a pound and those of us who measure calories each time we come near food.
As the author starts out:
The obese lack willpower; they overeat and underexercise — or so believe a majority of Americans. A 2012 online poll of 1143 adults conducted by Reuters and the market research firm Ipsos found that 61% of U.S. adults believed that “personal choices about eating and exercise” were responsible for the obesity epidemic. A majority of Americans, it seems, remain unaware of or unconvinced by scientific research suggesting that “personal choices” may not account for all cases of obesity.
No one doubts that there are a small few for whom weight control is highly problematic. No matter how low they go their Output is set well below that point. The few calories expended in exercise will never increase the output above the minimal input. But that does not account for the near global explosion of obesity. There was not a genetic change in humanity over the past thirty years to account for this. There is no epigenetic hyper/hypo methylation setting and resetting genes, causing massive reductions in basal Output. Input has increased by "choice" and all attempts to rationalize it as some effect out beyond human control is both wrong and harmful.
The law of mass balance applies. No one has ever rejected this. If it works in any chemical plant, and the human body is one, then it works in us. Remember the best meter is that scale and the best control mechanism is to not eat that extra candy bar or drink that extra soft drink. It is truly a shame that some physicians cannot call it for what it is. Oh, it appears as if the author is from the History Department at Harvard...
Net Neutrality and the FCC
The FCC issued its NPRM yesterday for new Net Neutrality. The rules are short, and the basis for substantial litigation.
Here is a sample of the key proposals:
§ 8.3 Transparency. (a) A person engaged in the provision of broadband Internet access service shall publicly disclose accurate information regarding the network management practices, performance, and commercial terms of its broadband Internet access services, in a manner tailored (i) for end users to make informed choices regarding use of such services, (ii) for edge providers to develop, market, and maintain Internet offerings, and (iii) for the Commission and members of the public to understand how such person complies with the requirements described in sections 8.5 and 8.7 of this chapter. (b) In making the disclosures required by this section, a person engaged in the provision of broadband Internet access service shall include meaningful information regarding the source, timing, speed, packet loss, and duration of congestion. (c) In making the disclosures required by this section, a person engaged in the provision of broadband Internet access service shall publicly disclose in a timely manner to end users, edge providers, and the Commission when they make changes to their network practices as well as any instances of blocking, throttling, and pay-for-priority arrangements, or the parameters of default or “best effort” service as distinct from any priority service.
Here is a sample of the key proposals:
§ 8.3 Transparency. (a) A person engaged in the provision of broadband Internet access service shall publicly disclose accurate information regarding the network management practices, performance, and commercial terms of its broadband Internet access services, in a manner tailored (i) for end users to make informed choices regarding use of such services, (ii) for edge providers to develop, market, and maintain Internet offerings, and (iii) for the Commission and members of the public to understand how such person complies with the requirements described in sections 8.5 and 8.7 of this chapter. (b) In making the disclosures required by this section, a person engaged in the provision of broadband Internet access service shall include meaningful information regarding the source, timing, speed, packet loss, and duration of congestion. (c) In making the disclosures required by this section, a person engaged in the provision of broadband Internet access service shall publicly disclose in a timely manner to end users, edge providers, and the Commission when they make changes to their network practices as well as any instances of blocking, throttling, and pay-for-priority arrangements, or the parameters of default or “best effort” service as distinct from any priority service.
The issue is what is to be disclosed. Congestion is a real difficult problem and it can be obfuscated readily. I have had difficulty adequately collecting such data for fifty years. There is no definition readily acceptable.
§ 8.5 No Blocking. A person engaged in the provision of fixed broadband Internet access service, insofar as such person is so engaged, shall not block lawful content, applications, services, or non-harmful devices, subject to reasonable network management. A person engaged in the provision of mobile broadband Internet access service, insofar as such person is so engaged, shall not block consumers from accessing lawful websites, subject to reasonable network management; nor shall such person block applications that compete with the provider’s voice or video telephony services, subject to reasonable network management.
§ 8.5 No Blocking. A person engaged in the provision of fixed broadband Internet access service, insofar as such person is so engaged, shall not block lawful content, applications, services, or non-harmful devices, subject to reasonable network management. A person engaged in the provision of mobile broadband Internet access service, insofar as such person is so engaged, shall not block consumers from accessing lawful websites, subject to reasonable network management; nor shall such person block applications that compete with the provider’s voice or video telephony services, subject to reasonable network management.
Wow, reasonable! That is how lawsuits are started. My reasonable and your reasonable are always at odds. This is clearly the most unrealistic document ever created!
§ 8.7 No Commercially Unreasonable Practices. A person engaged in the provision of fixed broadband Internet access service, insofar as such person is so engaged, shall not engage in commercially unreasonable practices. Reasonable network management shall not constitute a commercially unreasonable practice.
§ 8.7 No Commercially Unreasonable Practices. A person engaged in the provision of fixed broadband Internet access service, insofar as such person is so engaged, shall not engage in commercially unreasonable practices. Reasonable network management shall not constitute a commercially unreasonable practice.
If you liked the confusion above, then the last one noted here is even better! What does commercially unreasonable mean and to whom! This will be in the Courts for lifetimes! The big carriers could not be happier. I wonder who wrote this in the first place? Just a thought.
NOTCH, miR-146a and Melanoma
The understanding of cancer progression has been a
continuously evolving process moving from internal pathway elements, to micro
RNA interactions and to a complete set of epigenetic factors including various
methylation effects. We consider here a recent observation of a micro RNA
initiated by change in a pathway element, BRAF V600, and its effect on
proliferation and survival. This work by Forloni et al introduces putative new
elements to block with a putative therapeutic result, one in fact that handles
a multiplicity of factors.










In a recent paper by Forloni et al the authors state:
Oncogenic mutations in BRAF and NRAS occur in 70% of
melanomas. In this study, we identify a specific microRNA, miR-146a, which is
highly upregulated by oncogenic BRAF and NRAS. Expression of miR-146a increases
the ability of human melanoma cells to proliferate in culture and form tumors
in mice, whereas knockdown of miR-146a has the opposite effects. We show these
oncogenic activities are due to miR-146a targeting the NUMB mRNA, a repressor
of Notch signaling.
The focus is now clearly on these secondary factors, namely
the micro RNAs and even methylation effects that are seen in many cancers. In
this case it of the excess production of a specific miRNA that in turn block an
mRNA and in turn allows upregulation of other pathways and in turn unregulated
cell proliferation.
As Garraway states in NEJM:
Finally, these findings invite speculation that adding
γ-secretase inhibitors to inhibitors of RAF and MEK might offer an attractive
therapeutic cocktail for assessment in future clinical trials of melanoma
treatment. Given the substantial toxicity of γ-secretase inhibitors, additional
preclinical studies of such combinations in melanoma cell lines and patient
derived xenograft models would be beneficial.
Such studies could clarify the generalizability of Notch
dependency in melanoma, the relevance (if any) of the pre miR146a G allele
versus the C allele for patient stratification, and the possible usefulness of
alternative dosing and scheduling schema to reduce toxicity. Overall, this
study provides a reminder that, despite numerous advances, we have only just
begun to dissect the rich interplay among noncoding RNAs, the biologic basis of
cancer, and potential therapeutic strategies.
Garraway sees this as a significant breakthrough and believe
it is a worthwhile pathway for new therapeutics. In this analysis we briefly
examine the interaction of Notch and miR-146a and how it can be understood in
the case of melanoma as a major factor of uncontrolled proliferation.
Proteolysis is the process of degradation of proteins in the
cell and the release of the energy contained therein for other purposes. The
Notch pathway process is a key part of the proteolysis effort[1]. The
Notch system is a proteolytic driven system used in signal transduction in
cells. Uncontrolled Notch pathways production can lead to uncontrolled cellular
growth.
Let us begin with a simplified but reflective description of
the Notch pathway. The Notch process starts with the two Notch ligands, which
are also called DSL proteins. One is external to the cell membrane and is the
other is internal. When they are broken, the intracellular part, called NICD
moves to the nucleus and binds with a protein CSL which becomes a putative
transcription factor. We demonstrate that below.
Recall, that a transcription factor is a protein or protein
complex that can turn on (activators) or turn off (repressors) the transcription
of genes[2].
In this case the transcription factor is an activator for MYC[3].
Transcription factors are frequently brought to bear to activate genes that
lead to uncontrolled growth.
Goss and Kahn have presented a review of the interaction of
Notch and Wnt and especially the function of excess Notch activation as a part
of cell proliferation in multiple cancers[4].
As they state Wnt and Notch act in concert in many cancers, prostate being one
which we have examined in some detail. In addition excess Notch activation
appears to effect a stem cell like behavior in these cells thus resembling the
cell types enable for proliferation as well as survival.
We now want to explore some of the impacts of Notch in stem cell
environments and in turn in the maturation of cells. We focus on a recent paper
by Katoh and Katoh. As Katoh and Katoh have written:
Notch signaling pathway is implicated in the maintenance
of self-renewal potential in stem cells, binary cell-fate determination in
progenitor cells, and induction of terminal differentiation in proliferating
cells. Notch-ligand binding to Notch receptors leads to the cleavage of Notch
receptors by metalloprotease and Á-secretase to induce nuclear translocation of
Notch intracellular domain (NICD). Nuclear complex, consisting of CSL (RBPSUH),
NICD, Mastermind (MAML), p300 and histone acetyltransferase (HAT), then induces
transcriptional activation of Notch target genes, such as HES1, HES5, HES7,
HEY1, HEY2 and HEYL. HES/HEY family members are bHLH-type transcriptional
repressors for tissue-specific transcription factors. Therefore, Notch
signaling activation in stem cells leads to the maintenance of self-renewal
potential.
Now Katoh and Katoh provide an activation path progression
as show below (as modified):
The above demonstrates the progress from an overactive Notch
cell which thus acts as a stem cell to more mature cell lines. The above also
demonstrates the location of proliferating cells in this schema, just after the
stem cell line progenitor.
Thus the activation of Notch leads to an extreme survival
capability in cells so activated. They continue with the following regarding
NUMB:
NUMB and NUMB-like (NUMBL), consisting of phosphotyrosine-binding
(PTB) domain and SH3-binding proline-rich region, are docking proteins
functioning as Notch signaling inhibitors. Here, we searched for the
TCF/LEF-binding site within NUMB and NUMBL promoters. Because two
TCF/LEF-binding sites were identified within human NUMB promoter, comparative
integromics analyses on NUMB orthologs were further performed.
Thus one way to over-activate Notch is to suppress NUMB.
NUMB is described by NCBI as follows[5]:
The protein encoded by this gene plays a role in the
determination of cell fates during development. The encoded protein, whose
degradation is induced in a proteasome-dependent manner by MDM2, is a
membrane-bound protein that has been shown to associate with EPS15, LNX1, and
NOTCH1.
In a similar manner NOTCH1 is described as follows[6]:
This gene encodes a member of the Notch family. Members
of this Type 1 transmembrane protein family share structural characteristics
including an extracellular domain consisting of multiple epidermal growth
factor-like (EGF) repeats, and an intracellular domain consisting of multiple,
different domain types. Notch family members play a role in a variety of
developmental processes by controlling cell fate decisions.
The Notch signaling network is an evolutionarily
conserved intercellular signaling pathway which regulates interactions between
physically adjacent cells. …Homologues of the notch-ligands have also been
identified in human, but precise interactions between these ligands and the
human notch homologues remain to be determined. This protein is cleaved in the
trans-Golgi network, and presented on the cell surface as a heterodimer. This
protein functions as a receptor for membrane bound ligands, and may play
multiple roles during development.
These are two powerful and interacting genes. NUMB
suppresses Notch1 and Notch1 when activated makes for cell proliferation and
survival.
There are now well over hundreds of micro RNAs, which a
small non-coding RNAs which result in the control of various pathways in cellular
signalling. Micro RNAs are often encoded in introns in mRNAs and some in in
non-coding RNAs. They generally control mRNA in terms of its stability,
degradation and/or translation. The micro RNAs can stop genes from being
expressed as proteins, even though the gene is present and provides a normal
mRNA. They are small, generally 22 base pairs in length.
As NCBI states[7]:
microRNAs (miRNAs) are short (20-24 nt) non-coding RNAs
that are involved in post-transcriptional regulation of gene expression in
multicellular organisms by affecting both the stability and translation of
mRNAs. miRNAs are transcribed by RNA polymerase II as part of capped and
polyadenylated primary transcripts (pri-miRNAs) that can be either
protein-coding or non-coding.
The primary transcript is cleaved by the Drosha
ribonuclease III enzyme to produce an approximately 70-nt stem-loop precursor
miRNA (pre-miRNA), which is further cleaved by the cytoplasmic Dicer
ribonuclease to generate the mature miRNA and antisense miRNA star (miRNA*)
products. The mature miRNA is incorporated into a RNA-induced silencing complex
(RISC), which recognizes target mRNAs through imperfect base pairing with the
miRNA and most commonly results in translational inhibition or destabilization
of the target mRNA.
As Rusca and
Monticelli state:
Initial evidences on the possible involvement of miR-
146a in cancer came from a study showing thatmiR-146a was upregulated in
papillary thyroid carcinoma (PTC) samples compared with unaffected thyroid
tissue. Interestingly, a set of five miRNAs, including miR-221, miR-222, and
miR- 146, was sufficient to distinguish unequivocally between PTC and normal
thyroid. Similarly to the observations performed in immunologic settings,
overexpression of miR- 146a/b in the highly metastatic human breast cancer cell
line MDA-MB-231 significantly downregulated expression of IRAK1 and TRAF6,
negatively regulating NF-κB activity. Functionally, this resulted in markedly
impaired invasion and migration capacity relative to control cells.
These findings implicated miR-146 not only as a negative
regulator of constitutive NF-κB activity in breast cancer cells, but also
suggested that modulating miR-146 levels might have therapeutic potential to
suppress breast cancer metastases. Along the same line, miR-146a was among the
miRNAs found upregulated in cervical cancer tissues compared to normal cervix.
When introduced into cell lines, miR- 146a promoted cell
proliferation. Although the molecular mechanism underlying such increased
proliferation remains to be investigated, these observations potentially
implicate miR-146a in cervical carcinogenesis. In another type of cancer, the
hormone-refractory prostate carcinoma (HRPC), miR-146a levels were diminished
compared to androgen-sensitive noncancerous epithelium. In this context, miR-
146a acted as a tumor suppressor, reducing levels of its target ROCK1, one of
the key kinases involved in HRPC transformation.
Accordingly, forced miR-146a expression reduced ROCK1
protein levels, cell proliferation, invasion, and metastasis to human bone
marrow endothelial cell monolayers. Similarly, miR-146a was lower in pancreatic
cancer cells compared with normal human pancreatic cells…
There is now increasing evidence to suggest that miR-146a
is involved in the regulation of the adaptive as well as innate immune
response, and that miR-146a can be an important player in regulating tumor
progression.
However, more work remains to be done to fully understand
its role and mechanism of action in normal and pathologic conditions, so that
expression of this miRNA can potentially be exploited as a new point of entry
for therapy. With the identification of a vast number of miRNAs each carrying a
long list of putative targets, the challenge is now to understand the details
of their biological functions.
Thus miR-146a has significant roles to play in controlling
cell behavior.
For example, miRNAs can inhibit the translation of mRNA into
a protein. We show this below. The small segment attaches to the mRNA and
blocks translation. This graphic is descriptive and does not contain full
details[8].
In the case being discussed, miR-146a binds to NUMB and
suppresses it. That in turn allows for an overexpression of Notch which in turn
can lead to an unstable system with feedback. We shall detail that a bit later.
We depict that process in some detail below. For the most
part all miRNAs appear to function in the same manner. There are well over a
thousand identified at this point and more than likely many more to be found.
The functions of most are not fully known.
Before continuing it is worth a quick review of normal and
abnormal behavior of miRNA. The normal process is shown below. This shows a
classic blocking of translation. The miRNA binds to the mRNA and inhibits
translation. The question is what makes the miRNA to do this? Namely what
forces the generation of the miRNA? Is it a random effect or is it part of a
planned process. We have shown that homeostasis is well defined in terms of a
balanced expression of RNA. Yet when we have an aberrant genetic element the
miRNA can express itself in deleterious ways.
Now we can examine a miRNA in the context of a cancerous
environment shown below. The diagram below shows miRNA blocking a tumor
suppressor gene. In a sense the example of the miR-146a is an example of this
type of miRNA operation. It blocks a protein which in turn blocks a protein
which leads to unbridled growth and survival, Notch.
Finally we show the example of miRNA in some explosive
expansion of itself thus blocking many tumor suppressor genes. This is a deadly
mode for miRNAs and can be found in many cancers[9].
The classic pathway dynamics we understand regarding proliferation
and survival is shown below. This is the BRAF and PI3K dynamics. We demonstrate
this below. This is a well-known and well understood pathway and is at the core
of the BRAF V600 therapeutic approach.
Now proliferation and survival require gene activation and
maintenance.
In this report, we demonstrate a critical role for
miR-146a in the initiation and progression of BRAF/ NRAS-positive melanomas,
... In addition, our results reveal a pharmacologically tractable pathway for
the treatment of melanoma. We identified miR-146a as the microRNA whose
expression was most upregulated by activated BRAF.
Upregulation of miR-146a by activated BRAF, as well as
activated NRAS, occurs through the MAPK signaling pathway. Accordingly, we find
that BRAF and NRAS mutant melanoma cell lines and short-term melanoma cultures
show higher levels of miR-146a compared to those that are wild type for these
genes.
A major function of the MAPK pathway is to activate
transcription by regulating the stability and expression of multiple
transcription factors primarily through direct phosphorylation.
We show that the MAPK pathway regulates the
phosphorylation of the transcription factor MYC, which in turn binds to the
promoter of miR-146a and stimulates its transcription. Notably, MYC has been
found to stimulate transcription of several other miRNAs. For example, MYC has
been shown to directly activate transcription of the oncogenic miR-17-92
cluster and thereby promote cell proliferation, survival, angiogenesis, and
metabolic reprogramming in a number of tumor cell lines.
miRNAs and components of miRNA biogenesis pathways such
as Dicer have been implicated in several aspects of melanocyte biology as well
as in melanoma initiation and progression.
We depict some of that process below:
Previous studies
have shown that miR-146a can function either as an oncogene or as a tumor
suppressor depending upon the cell type. For example, miR-146a has been shown to
function as an oncogene in a variety of human cancers including papillary
thyroid carcinoma (PTC), triple negative sporadic breast cancers and anaplastic
thyroid carcinoma. miRNAs function primarily by targeting mRNAs and either
promoting their degradation or blocking their translation.
Our analysis identified 20 potential targets of miR-146a,
including NUMB, which is a well-characterized Notch signaling inhibitor. It is
thought that NUMB negatively regulates NOTCH, potentially through a direct
protein–protein interaction that requires the phosphotyrosine-binding (PTB)
domain of NUMB and either the RAM23 region or the very C-terminal end of NOTCH.
We demonstrate some of these dynamics in the Figure below
(adapted from Galloway with modifications).
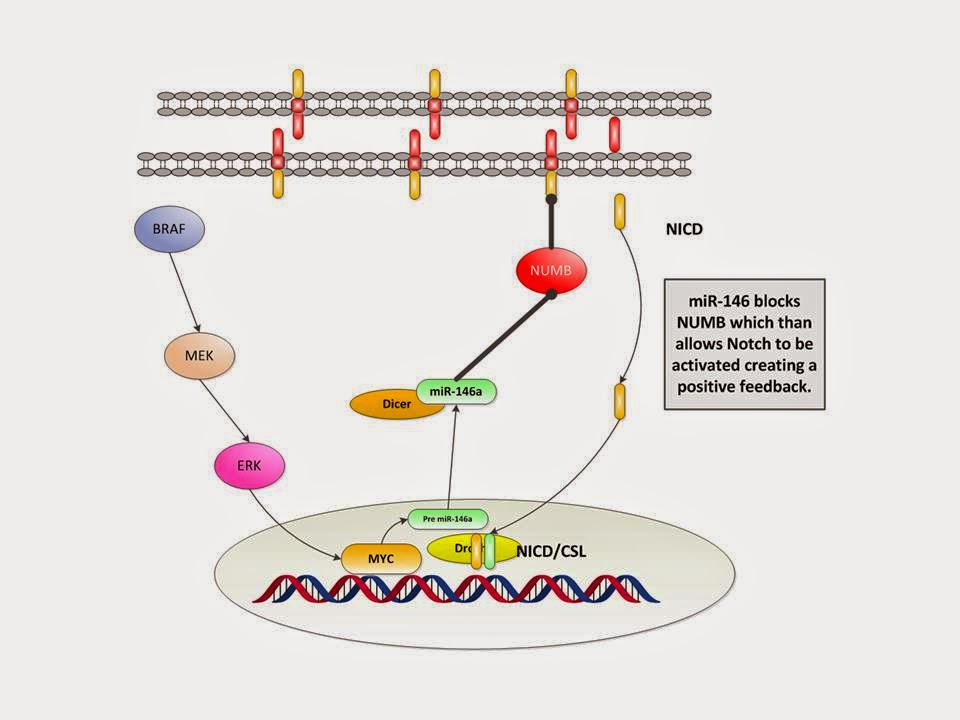
In simple terms:
1. BRAF activates MEK
2. and then ERK
3. which activates MYC
4. which activates pre miR-146a and
5. then via Drosha and Dicer makes miR-146a
6. which reactivates Notch by suppressing NUMB expression (we have
left that out for simplicity)
7. which then goes down to the transcription on the DNA resulting
in proliferation and stem like behavior.
This is an interesting and compelling mechanism for the
explanation of the aggressive melanoma expansion.
This is an interesting step in the understanding of melanoma
genomics. The role of micro RNAs is becoming clearer as time goes by and added
to that is the effect of such epigenetic factors as methylation and we now see
a much more complex field of play than a decade ago. The benefit is the
recognition of more targets of opportunity that can be had for potential
therapeutics. On the other hand the main concern is that the more that is
learned one may ask what else is there yet to grasp.
Thus what observations can we make here? Let us examine a
few:
1. Stem Cell Hypothesis. Here we have the elements of how a
stem cell functions with the activated Notch and blocked NUMB. Does this imply
that we have the re-emergence of stem cell like malignant cells activated in a
manner such as this. Namely the miRNAs allow for the reprogramming of some
modified form of totipotency.
2. Targeted Therapeutics: Galloway makes this observation We
know that BRAF inhibitors get us one step there but then we need MEK
inhibitors. Then what? Does an inhibitor for miR-146a take all the steps
necessary or does the cell go and find another back door way to function?
3. The Dynamics of the Processes are Not Well Understood.
One of the problems in understanding the impact of miRNAs and other pathway
elements is that there is a concern as to the number or concentration of
products. If miR-146a is to block NUMB then it should block all NUMB and in
turn activate all Notch. Yet it is a molecule by molecule process which seems
to be poorly understood. The paper by Choir et al and Nazarov et al present
some ideas on how to deal with such issues. However they are but first steps.
This is a critical factor to understand since the therapeutics depends on
blocking the necessary number of miR-146a molecules. To data there seems to be
limited data to assess this issue.
4. Initiation and Support: We know that V600 mutation of
BRAF is drivers for metastatic melanoma. However it is not clear what is the
driver ultimately for miR-146a, although it appears as we have suggested as a
sequella from the other mutations. Additional insight into the proliferation is
requires.
1.
Appasani, K., MicroRNAs,
Cambridge (New York) 2008.
2.
Broad Institute, https://www.broadinstitute.org/education/glossary
3.
Choi, M., et al, A dynamic
expression survey identifies transcription factors relevant in mouse digestive
tract development, Development 133, 4119-4129 (2006) doi:10.1242/dev.02537
4.
Forloni, M., et al,
miR-146a promotes the initiation and progression of melanoma by activating
Notch signaling, eLife 2014;3: e01460. DOI: 10.7554/eLife.01460
5.
Garraway, L., A Notch for
Noncoding RNA in Melanoma, NEJM, 370;20, May 15, 2014.
6.
Goss, K., M. Kahn,
Targeting the Wnt Pathway in Cancer, Springer (New York) 2011.
7.
Katoh, M., M., Katoh, NUMB
is a break of WNT - Notch signaling cycle, INTERNATIONAL JOURNAL OF MOLECULAR
MEDICINE 18: 517-521, 2006
8.
Lawrie, C., MicroRNAs in
Medicine, Wiley (Hoboken) 2014.
9.
Marks, F., et al, Cellular
Signal Processing, Garland 2008.
10.
McGarty, T., Melanoma
Genomics, DRAFT 2013.
11.
McGarty, T., Prostate
Cancer Genomics, DRAFT, 2013.
12.
Nazarov, P., et al,
Interplay of microRNAs, transcription factors and target genes: linking dynamic
expression changes to function, Nucleic Acids Research, 2013, Vol. 41, No. 5
2817–2831.
13.
Rusca, S., C. Monticelli,
MiR-146a in Immunity and Disease, Molecular Biology International, Volume 2011,
Article ID 437301, 7 pages
14.
Sangunitti, et al,
Probabilistic inference of transcription factor concentrations and
gene-specific regulatory activities, Bioinformatics, Vol. 22 no. 22 2006, pages
2775–2781
15.
Vaquerizas, J., et al, A
census of human transcription factors: function, expression and evolution,
Nature Reviews Genetics 10, 252-263 (April 2009) | doi:10.1038/nrg2538.
16.
Watson, J., et al,
Molecular Biology of the Gene, 5th Ed, Benjamin (San Francisco)
2004.
[1]
See Marks, Chapter 13.
[2]
See Broad https://www.broadinstitute.org/education/glossary/transcription-factor
and Watson et al 544-555. Also http://www.nature.com/scitable/definition/general-transcription-factor-transcription-factor-167 Also from Vaquerizas:
“Transcription factors are key cellular components
that control gene expression: their activities determine how cells function and
respond to the environment. Currently, there is great interest in research into
human transcriptional regulation. However, surprisingly little is known about
these regulators themselves. For example, how many transcription factors does
the human genome contain? How are they expressed in different tissues? Are they
evolutionarily conserved? Here, we present an analysis of 1,391 manually
curated sequence-specific DNA-binding transcription factors, their functions,
genomic organization and evolutionary conservation. Much remains to be
explored, but this study provides a solid foundation for future investigations
to elucidate regulatory mechanisms underlying diverse mammalian biological
processes.”
[3]
From NCBI we have: The protein encoded by this gene, cMYC, is a
multifunctional, nuclear phosphoprotein that plays a role in cell cycle
progression, apoptosis and cellular transformation. It functions as a
transcription factor that regulates transcription of specific target genes. Mutations,
overexpression, rearrangement and translocation of this gene have been
associated with a variety of hematopoietic tumors, leukemias and lymphomas,
including Burkitt lymphoma. There is evidence to show that alternative
translation initiations from an upstream, in-frame non-AUG (CUG) and a
downstream AUG start site result in the production of two isoforms with
distinct N-termini. The synthesis of non-AUG initiated protein is suppressed in
Burkitt's lymphomas, suggesting its importance in the normal function of this
gene. See http://www.ncbi.nlm.nih.gov/gene/4609
[4]
See Goss and Kahn, pp60-61.
[8]
See Marks p 318. Note the colors are also descriptive and do not reflect any
specific RNA base pair pairing. Just as with DNA we would expect similar
bonding of CG and A and U.
[9] It
is worth examining the McGarty DRAFTs on Prostate Cancer and Melanoma to see
this in some detail.
Thursday, May 15, 2014
Cable Unbundling Illogic
There is a piece today in the NY Times which rambles about why unbundling cable channels will cost us all more. Stop right there. It will cost everyone more, more than the extortion already in place. Again, I have never seen a football game, one baseball game, one basketball game, one hockey game, really just half, and that was almost fifty years ago. Yet I pay almost $20 a month in cable charges for all the stuff I have no interest in. $70 a month for basic cable! If I get less then I pay more? Huh?
The article starts:
If you have cable TV, you probably don’t watch most of the channels you get. The average American television household receives 189 channels, up from 123 in 2003. But we’re watching only 17.5 of those channels — nearly unchanged from 11 years ago, according to a new report from Nielsen.
I do not know what those channels are but I damn well pay for them. Now try this on for size:
Think of it this way: If I put my bag in an overhead luggage bin, you can’t put your bag in the same spot, so it makes sense to charge me personally for my use. But if I watch Bravo, that doesn’t stop anyone else from watching the same show. When a good is “nonrivalrous” like a cable signal, giving it to me doesn’t stop anyone else from using it or add production costs at the margin. In those cases, it can make sense to throw lots of stuff into one package, whether or not I’ll actually use it.
The fact is that they make money from advertising and for the forcible fees I pay whether I watch Duck Dynasty or not. Advertising is based on reach, measures of real customers watching. So if it were all advertising then there is no need for me to watch unless I find it of some value. No value, then I do not watch and do not buy. However the cable guys contort all logic. What is the guy saying in the above. I only speak six languages and the words above are unrecognizable.
The author also uses the argument that one bundles a newspaper. But simply one can readily unbundle the electronic content over a cable. One has more difficulty in doing so in print! In fact, if one priced cable as access plus content then content would become truly market based. It would engender true competition. As it stands now one can bundle absolute nonsense and get paid for it.
We do not argue about some bundling. It is the sports bundling that pays exorbitant salaries to athletes many of whom often get themselves into legal messes. No extortionary charges, unbundle the sports channels and reduce the salaries. Try Income Inequality on those folks for a change, Piketty, where are you when we really need you?
The article starts:
If you have cable TV, you probably don’t watch most of the channels you get. The average American television household receives 189 channels, up from 123 in 2003. But we’re watching only 17.5 of those channels — nearly unchanged from 11 years ago, according to a new report from Nielsen.
I do not know what those channels are but I damn well pay for them. Now try this on for size:
Think of it this way: If I put my bag in an overhead luggage bin, you can’t put your bag in the same spot, so it makes sense to charge me personally for my use. But if I watch Bravo, that doesn’t stop anyone else from watching the same show. When a good is “nonrivalrous” like a cable signal, giving it to me doesn’t stop anyone else from using it or add production costs at the margin. In those cases, it can make sense to throw lots of stuff into one package, whether or not I’ll actually use it.
The fact is that they make money from advertising and for the forcible fees I pay whether I watch Duck Dynasty or not. Advertising is based on reach, measures of real customers watching. So if it were all advertising then there is no need for me to watch unless I find it of some value. No value, then I do not watch and do not buy. However the cable guys contort all logic. What is the guy saying in the above. I only speak six languages and the words above are unrecognizable.
The author also uses the argument that one bundles a newspaper. But simply one can readily unbundle the electronic content over a cable. One has more difficulty in doing so in print! In fact, if one priced cable as access plus content then content would become truly market based. It would engender true competition. As it stands now one can bundle absolute nonsense and get paid for it.
We do not argue about some bundling. It is the sports bundling that pays exorbitant salaries to athletes many of whom often get themselves into legal messes. No extortionary charges, unbundle the sports channels and reduce the salaries. Try Income Inequality on those folks for a change, Piketty, where are you when we really need you?
Have You Ever Tried to Find Anything at Microsoft!
Now when one learns to use Microsoft products there is often the tendency by the company to change them to some new incomprehensible form. Just look at Windows 8. So when I saw this in PC World I though it is worth a comment.
Let’s face it—navigating through a maze of menus isn’t what any user wants to do. And that’s why Microsoft is moving its intelligent search box, Tell Me, from Word Online to similar versions of PowerPoint and Excel. Tell Me, fortunately, is a very simple concept: It’s a natural language search box, similar to what you might find in Microsoft’s Bing search engine or Google. In Word, Excel and PowerPoint Online, it sits at the top center of the Ribbon, with the helpful cue “Tell me what you want to do.” Simply type in your request, and it’s off to the races. (As a handy shortcut, you can even type CTRL-‘ (the apostrophe) to auto-place your cursor into the Tell Me field.
As anyone who has ever tried to figure out how to work some of Microsoft's issues off of their web site knows, "Abandon all hope ye who enter", Dante's Inferno is an amusement park compared to Microsoft Help. Go to Google, get the advice or input and then hack your way through Microsoft.
Office 2013 looks like a Kindergarten colored room, whomever chose the colors, well, no matter.
But really, use an online service to type, and then use Microsoft for help! Then use Bing! Now really guys, who put the magazine up to this one. Use Google, it works.
Let’s face it—navigating through a maze of menus isn’t what any user wants to do. And that’s why Microsoft is moving its intelligent search box, Tell Me, from Word Online to similar versions of PowerPoint and Excel. Tell Me, fortunately, is a very simple concept: It’s a natural language search box, similar to what you might find in Microsoft’s Bing search engine or Google. In Word, Excel and PowerPoint Online, it sits at the top center of the Ribbon, with the helpful cue “Tell me what you want to do.” Simply type in your request, and it’s off to the races. (As a handy shortcut, you can even type CTRL-‘ (the apostrophe) to auto-place your cursor into the Tell Me field.
As anyone who has ever tried to figure out how to work some of Microsoft's issues off of their web site knows, "Abandon all hope ye who enter", Dante's Inferno is an amusement park compared to Microsoft Help. Go to Google, get the advice or input and then hack your way through Microsoft.
Office 2013 looks like a Kindergarten colored room, whomever chose the colors, well, no matter.
But really, use an online service to type, and then use Microsoft for help! Then use Bing! Now really guys, who put the magazine up to this one. Use Google, it works.
Wednesday, May 14, 2014
More on Piketty
The splash of Piketty's thoughts on the evil of wealth, or income inequality, has led to many commentators saying one thing or another. Now in comes Summers with his thoughts. (I had written an analysis of Piketty and Francis that may be used as an alternate view.)
He begins with statements like the following:
This should not be surprising. At a moment when our politics seem to be defined by a surly middle class and the President has made inequality his central economic issue, how could a book documenting the pervasive and increasing concentration of wealth and income among the top 1, .1, and .01 percent of households not attract great attention? Especially when it exudes erudition from each of its nearly 700 pages, drips with literary references, and goes on to propose easily understood laws of capitalism that suggest that the trend toward greater concentration is inherent in the market system and will persist absent the adoption of radical new tax policies.
What surly middle class is he talking about? Yes the current President still acts as a community rabble rouser but that should be no surprise. But erudition. How many time do we need to be reminded of Balzac! I get the point. But if one were truly erudite then one would understand that Balzac in context was protesting his own social ills. Why not the mad man Dickens. But alas in the US we do not have dynasties, except attempts at in in politics. In my opinion the book is a gross bore with multiple repetitions of the same thing.
He does make a significant point:
A brief look at the Forbes 400 list also provides only limited support for Piketty’s ideas that fortunes are patiently accumulated through reinvestment. When Forbes compared its list of the wealthiest Americans in 1982 and 2012, it found that less than one tenth of the 1982 list was still on the list in 2012, despite the fact that a significant majority of members of the 1982 list would have qualified for the 2012 list if they had accumulated wealth at a real rate of even 4 percent a year. They did not, given pressures to spend, donate, or misinvest their wealth. In a similar vein, the data also indicate, contra Piketty, that the share of the Forbes 400 who inherited their wealth is in sharp decline.
Namely we do not have old style multi century European wealth. We have American entrepreneurial wealth. High risk and high return.
He then states:
Hanging over this subject is a last issue. Why is inequality so great a concern? Is it because of the adverse consequences of great fortunes or because of the hope that middle-class incomes could grow again? If, as I believe, envy is a much less important reason for concern than lost opportunity, great emphasis should shift to policies that promote bottom-up growth. At a moment when secular stagnation is a real risk, such policies may include substantially increased public investment and better training for young people and retraining for displaced workers, as well as measures to reduce barriers to private investment in spheres like energy production, where substantial job creation is possible.
That is the question I always start with. Why? Why should we worry about this issue?
The he states near the end:
Look at Kennedy airport. It is an embarrassment as an entry point to the leading city in the leading country in the world. The wealthiest, by flying privately, largely escape its depredations. Fixing it would employ substantial numbers of people who work with their hands and provide a significant stimulus to employment and growth. As I’ve written previously, if a moment when the United States can borrow at lower than 3 percent in a currency we print ourselves, and when the unemployment rate for construction workers hovers above 10 percent, is not the right moment to do it, when will that moment come?
Has have ever been to Penn Station? It makes Mumbai look like a palace. The answer is Politics! The Port Authority often has control and its management is filled with in my opinion pure political hacks. Bridges rust, tolls explode, and salaries increase. Look at Kennedy, look at Penn Station, look at Port Authority Bus Station and you see what happens with politicians emptying the pockets of the taxpayers with agencies reportable to no one! Give them more money, then it ends in the pockets of the Unions. This is a no win game.
He begins with statements like the following:
This should not be surprising. At a moment when our politics seem to be defined by a surly middle class and the President has made inequality his central economic issue, how could a book documenting the pervasive and increasing concentration of wealth and income among the top 1, .1, and .01 percent of households not attract great attention? Especially when it exudes erudition from each of its nearly 700 pages, drips with literary references, and goes on to propose easily understood laws of capitalism that suggest that the trend toward greater concentration is inherent in the market system and will persist absent the adoption of radical new tax policies.
What surly middle class is he talking about? Yes the current President still acts as a community rabble rouser but that should be no surprise. But erudition. How many time do we need to be reminded of Balzac! I get the point. But if one were truly erudite then one would understand that Balzac in context was protesting his own social ills. Why not the mad man Dickens. But alas in the US we do not have dynasties, except attempts at in in politics. In my opinion the book is a gross bore with multiple repetitions of the same thing.
He does make a significant point:
A brief look at the Forbes 400 list also provides only limited support for Piketty’s ideas that fortunes are patiently accumulated through reinvestment. When Forbes compared its list of the wealthiest Americans in 1982 and 2012, it found that less than one tenth of the 1982 list was still on the list in 2012, despite the fact that a significant majority of members of the 1982 list would have qualified for the 2012 list if they had accumulated wealth at a real rate of even 4 percent a year. They did not, given pressures to spend, donate, or misinvest their wealth. In a similar vein, the data also indicate, contra Piketty, that the share of the Forbes 400 who inherited their wealth is in sharp decline.
Namely we do not have old style multi century European wealth. We have American entrepreneurial wealth. High risk and high return.
He then states:
Hanging over this subject is a last issue. Why is inequality so great a concern? Is it because of the adverse consequences of great fortunes or because of the hope that middle-class incomes could grow again? If, as I believe, envy is a much less important reason for concern than lost opportunity, great emphasis should shift to policies that promote bottom-up growth. At a moment when secular stagnation is a real risk, such policies may include substantially increased public investment and better training for young people and retraining for displaced workers, as well as measures to reduce barriers to private investment in spheres like energy production, where substantial job creation is possible.
That is the question I always start with. Why? Why should we worry about this issue?
The he states near the end:
Look at Kennedy airport. It is an embarrassment as an entry point to the leading city in the leading country in the world. The wealthiest, by flying privately, largely escape its depredations. Fixing it would employ substantial numbers of people who work with their hands and provide a significant stimulus to employment and growth. As I’ve written previously, if a moment when the United States can borrow at lower than 3 percent in a currency we print ourselves, and when the unemployment rate for construction workers hovers above 10 percent, is not the right moment to do it, when will that moment come?
Has have ever been to Penn Station? It makes Mumbai look like a palace. The answer is Politics! The Port Authority often has control and its management is filled with in my opinion pure political hacks. Bridges rust, tolls explode, and salaries increase. Look at Kennedy, look at Penn Station, look at Port Authority Bus Station and you see what happens with politicians emptying the pockets of the taxpayers with agencies reportable to no one! Give them more money, then it ends in the pockets of the Unions. This is a no win game.
Subscribe to:
Posts (Atom)


